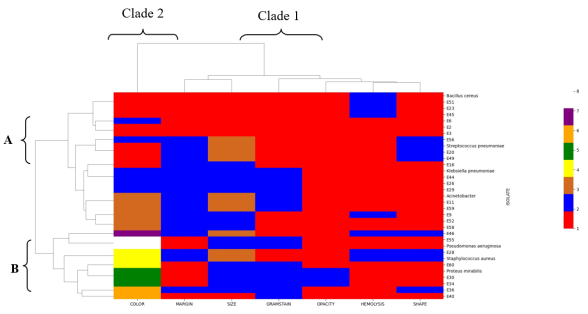
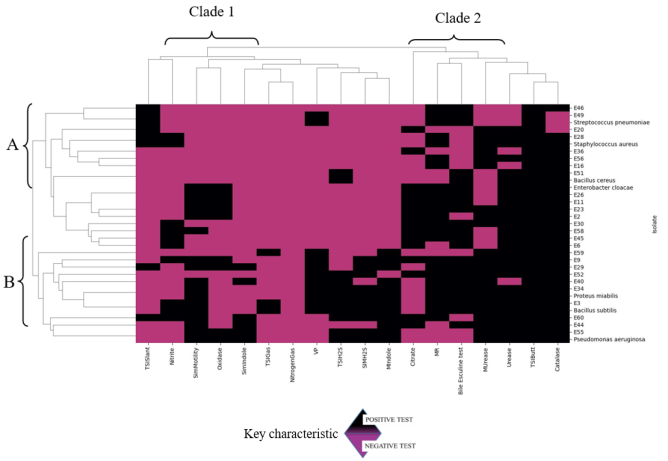
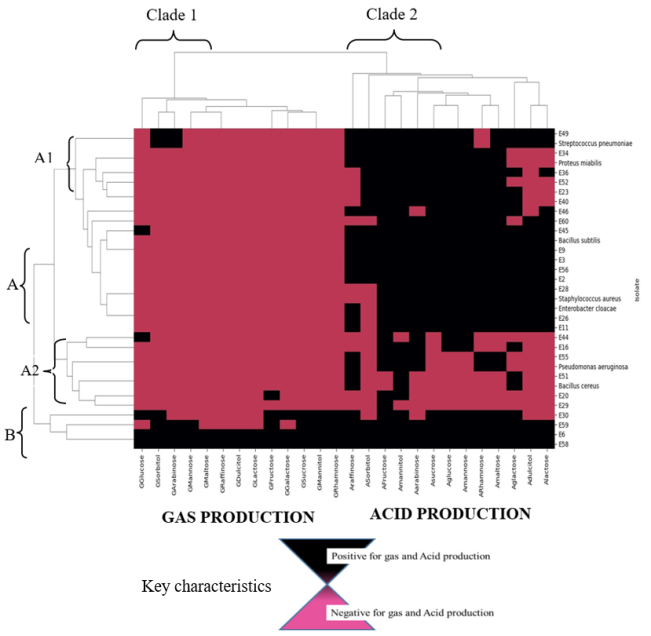
Facemasks have been widely used in the theater to prevent surgical site infections as well as in hospital wards to prevent infectious infections. Also masks have been recommended to be used by the public during pandemics of respiratory infectious diseases. However, the prolonged use of these masks may have inadvertently harbored unseen dangers by serving as reservoirs for bacteria contamination and antibiotic resistant bacteria when in use. The objective of this study was to quantify bacteria as well as isolate, characterize and determine the antibiotic profiles of bacteria from worn reusable facemasks. Before masks were worn, the participants’ mouth, nose and facial skin were swabbed with sterile transport media. Thereafter, the masks were worn at two-, four- and six-hour intervals. Afterwards, bacteria on worn masks, mouth, nose and skin were identified via morphological, biochemical and molecular methods. Antibacterial susceptibility was determined via the Kirby–Bauer method. The results of this study revealed bacterial colony forming units were significantly higher in the polycotton masks (4.30×102 CFUs) than in the cotton masks (3.38×102 CFUs). Colony forming units increased with extended mask use from two, four, and six hours respectively (7.61×101 CFUs, 1.28×102 CFUs, 4.26×102 CFUs). Also, inside part of the masks had significantly high colony forming units than outside of the masks. The isolated bacteria were from different genera including Bacillus, Staphylococcus, Enterococcus, Pseudomonas, Stenotrophomonas, Acinetobacter, Neisseria, Proteus, Klebsiella and Enterobacter. From the antibiotic resistance profiles, with 85.2% of isolates classified as resistant, Ampicillin had the highest resistance rate. Spectinomycin and Amoxicillin both showed 66.7% resistance, whereas Cefotaxime showed 63.0% resistance. Streptomycin showed the highest susceptibility (48.1%), suggesting comparatively improved efficiency against the isolates. The study findings provides bacteriological insights risks associated with facemask upon use as well as antibiotic resistance.
| Published in | International Journal of Food Science and Biotechnology (Volume 10, Issue 2) |
| DOI | 10.11648/j.ijfsb.20251002.12 |
| Page(s) | 33-48 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Bacteria, Reusable Mask (RM), Cotton Mask (CM), Polycotton Mask (PM), Antibiotic Resistance
Mask Types | Colony Forming Units (CFUs)/0.1 ml of 50 ml wash |
|---|---|
Cotton | 338.73±32.68b |
Poly-cotton | 430.28±47.75a |
Cotton control | 31.11±0.09c |
Poly-cotton control | 42±1.07c |
Time | |
2 hours | 76.12 ±5.33c |
4 hours | 128.56±9.09b |
6 hours | 426.92± 43.88a |
Location | |
Inside (face side) | 240.98± 26.39a |
Outside | 180.08±20.49b |
P values of the main effect and their interaction | |
Mask Type | <.0001 |
Time | <.0001 |
Location | 0.0013 |
Mask type*location | 0.0009 |
Location*Time | 0.0319 |
Mask type*Time | <.0001 |
Mask type*Location*Time | 0.1269 |
Antibiotic | % Susceptible (S) | % Intermediate (I) | % Resistant (R) |
|---|---|---|---|
Ampicillin | 14.8% | 0.0% | 85.2% |
Streptomycin | 48.1% | 3.7% | 48.1% |
Spectinomycin | 22.2% | 11.1% | 66.7% |
Cefotaxime | 22.2% | 14.8% | 63.0% |
Amoxicillin | 22.2% | 11.1% | 66.7% |
Sulfan | 29.6% | 7.4% | 63.0% |
COVID-19 | Coronavirus Disease 2019 |
SARS-CoV 2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
WHO | World Health Organization |
CFU | Colony Forming Unit |
RM | Reusable mask |
CM | Cotton mask |
PM | Polycotton mask |
TSI | Tripple Sugar Iron |
SIM | Sulfide Indole Motility |
MIU | Motility Indole Urease |
MR | Methyl Red |
VP | Voges–Proskauer |
DNA | Deoxyribonucleic acid |
CTAB | Cetrimonium bromide |
TBE | Tris-borate-EDTA |
UV | Ultraviolet radiation |
ANOVA | Analysis of Variance |
SAS | Statistical Analysis Software |
BLAST | Basic Local Alignment Search Tool |
NCBI | National Council of Biotechnology Institute |
MEGA-X | Molecular Evolutionary Genetics Analysis across Computing Platforms. |
HSD | Tukey’s Honest Significance Difference |
PCR | Polymerase Chain Reaction |
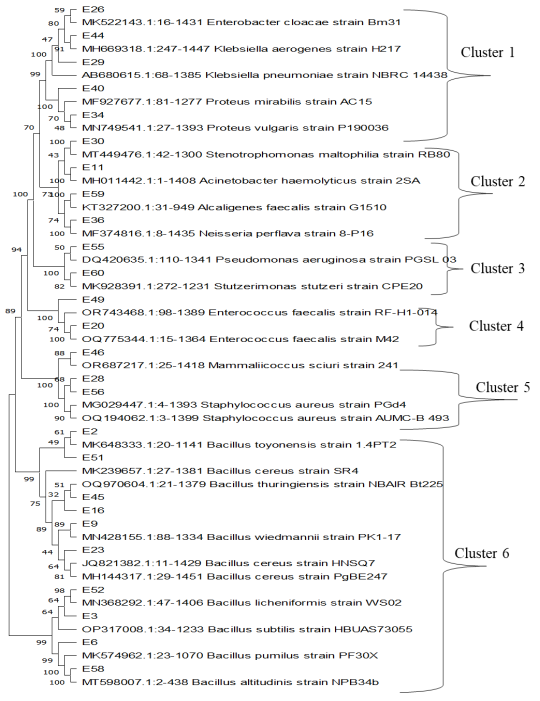
Isolate | Source | Identity | Percentage Identity | Accession Number |
|---|---|---|---|---|
E2 | CO, PI | Bacillus toyonensis | 94.39 | PP408994 |
E3 | CO, M, CI | Bacillus subtilis | 96.26 | PP408970 |
E6 | PI, CI | Bacillus pumillus | 95.90 | PP408991 |
E9 | CI | Bacillus wiedmannii | 92.36 | PP408978 |
E11 | CI, M | Acinetobacter hemoolyticus | 98.23 | PP408988 |
E16 | PO, CI | Bacillus thuringiensis | 98.24 | PP408972 |
E20 | PI, CI, M | Enterococcus faecalis | 99.11 | PP408973 |
E23 | PI, CI, S, N | Bacillus cereus | 99.51 | PP408976 |
E26 | PI | Enterobacter cloacae | 97.16 | PP408989 |
E28 | CI, CO, PI, PO, S, M, N | Staphylococcus aureus | 99.21 | PP408975 |
E29 | C1, M | Klebsiella pneumoniae | 98.79 | PP408979 |
E30 | N | Stenotrophomonas maltophilia | 98.73 | PP408974 |
E34 | PI | Proteus mirabilis | 98.25 | PP408971 |
E36 | PI, CI, S, M | Neisseria perflava | 92.33 | PP408987 |
E40 | CO | Proteus vulgaris | 90.43 | PP408980 |
E44 | M | Klebsiella aerogenes | 90.43 | PP408990 |
E45 | PI, CI, S, M, N | Bacillus cereus | 99.51 | PP408981 |
E46 | PI, CI, M, N | Mammliicoccus sciurii | 95.63 | PP408982 |
E49 | PI, CO, S, M | Enterococcus faecalis | 99.69 | PP408986 |
E51 | CO | Bacillus cereus | 96.25 | PP408977 |
E52 | PI, CI, S, N | Bacillus licheniformis | 94.64 | PP408985 |
E55 | PI, M | Pseudomonas aeruginosa | 86.79 | PP408984 |
E56 | PI, CI, S, M, N | Staphylococcus aureus | 96.14 | PP408983 |
E58 | CO, S, N | Bacillus altitudinis | 100 | PP437268 |
E59 | PI | Alcaligenes faecalis | 100 | PP408993 |
E60 | PO | Stutzerimonas stutzeri | 100 | PP408992 |
ISOLATE | ZONES OF INHIBITION(Millimeter) mm | |||||
|---|---|---|---|---|---|---|
Ampicillin | Streptomycin | Spectinomycin | Cefotaxime | Amoxicillin | Sulfan | |
E2 | 6.00±0.00g | 24.00±0.58cde | 25.67±0.33b | 6.00±0.00j | 14.00±0.00fgh | 6.00±0.00e |
E3 | 9.00±0.00e | 10.00±0.00ij | 14.00±0.00fgh | 8.00±0.00ij | 35.67±2.60a | 37.00±2.89a |
E6 | 9.00±0.00e | 28.67±0.88a | 8.00±0.00jk | 19.00±0.58d | 22.00±1.15de | 13.00±0.58d |
E9 | 6.00±0.00g | 28.00±1.15ab | 24.00±0.57bc | 11.00±0.00gh | 13.67±1.45fgh | 6.00±0.00e |
E11 | 13.33±0.67d | 8.00±0.00jk | 10.00±0.00ij | 22.00±0.00c | 12.00±0.00ghi | 18.00±0.00c |
E16 | 6.00±0.00g | 6.00±0.00k | 10.33±0.88i | 29.67±0.33b | 14.00±0.00fgh | 6.00±0.00e |
E20 | 7.00±0.00fg | 8.00±0.00jk | 6.00±0.00k | 16.00±1.15e | 6.00±0.00k | 8.00±0.00e |
E23 | 7.00±0.00fg | 6.00±0.00k | 6.00±0.00k | 8.00±0.00ij | 6.00±0.00k | 6.00±0.00e |
E26 | 6.00±0.00g | 6.00±0.00k | 8.00±0.00jk | 11.00±0.00gh | 7.00±0.00jk | 12.00±0.00d |
E28 | 6.00±0.00g | 25.00±0.00cd | 30.00±0.00a | 6.00±0.00j | 18.00±0.00ef | 24.00±0.00b |
E29 | 14.00±0.00d | 12.00±0.00i | 22.00±0.00cd | 10.00±0.00ghi | 12.00±0.00ghi | 18.00±0.00c |
E30 | 8.00±0.00ef | 28.00±0.00ab | 6.00±0.00k | 14.00±0.00ef | 8.00±0.00ijk | 14.00±0.00d |
E34 | 6.00±0.00g | 30.00±0.00a | 15.67±0.33efg | 32.00±1.15b | 11.00±0.00hij | 6.00±0.00e |
E36 | 8.67±0.33e | 6.00±0.00k | 13.67±0.88gh | 6.00±0.00j | 6.00±0.00k | 23.00±0.58b |
E40 | 6.00±0.00g | 19.67±0.33g | 17.00±0.00e | 20.33±0.88cd | 12.67±0.33gh | 8.00±0.00e |
E44 | 6.00±0.00g | 16.00±0.00h | 10.00±0.00ij | 6.00±0.00j | 16.00±0.00fg | 14.00±0.00d |
E45 | 6.00±0.00g | 7.00±0.00k | 6.00±0.00k | 9.00±0.00hi | 6.00±0.00k | 6.00±0.00e |
E46 | 20.00±0.00c | 20.00±0.00gf | 11.00±0.58i | 22.00±1.15c | 29.00±0.58bc | 26.00±0.00b |
E49 | 8.00±0.00ef | 6.00±0.00k | 6.00±0.00k | 11.67±0.33fg | 7.00±0.00jk | 6.00±0.00e |
E51 | 6.00±0.00 g | 23.67±0.88de | 12.00±0.00hi | 6.00±0.00j | 24.00±0.88cd | 14.67±0.88cd |
E52 | 6.00±0.00g | 6.00±0.00k | 20.00±0.00d | 6.00±0.00j | 6.00±0.00k | 6.00±0.00e |
E55 | 6.00±0.00g | 10.00±0.00ij | 6.00±0.00k | 18.67±0.33d | 6.00±0.00k | 6.00±0.00e |
E56 | 23.00±0.00b | 22.00±0.58cd | 6.00±0.00k | 8.00±0.00ij | 18.00±0.00ef | 35.00±0.00a |
E58 | 32.00±0.58a | 25.00±0.58cd | 28.00±1.15a | 6.00±0.00j | 36.00±2.31a | 38.00±0.00a |
E59 | 6.00±0.00g | 26.00±0.58bc | 6.00±0.00k | 15.00±0.00e | 6.00±0.00k | 6.00±0.00e |
E60 | 20.00±0.00c | 23.00±0.00de | 16.00±0.58ef | 35.00±0.00a | 33.00±0.58ab | 23.00±0.58b |
Controls | 6.00±0.00g | 6.00±0.00k | 6.00±0.00k | 6.00±0.00j | 6.00±0.00k | 6.00±0.00e |
P values | <.0001 | <.0001 | <.0001 | <.0001 | <.0001 | <.0001 |
Isolate | Ampicillin | Streptomycin | Spectinomycin | Cefotaxime | Amoxicillin | Sulfan |
|---|---|---|---|---|---|---|
E2 | R | S | S | R | R | R |
E3 | R | R | R | R | S | S |
E6 | R | S | R | I | S | R |
E9 | R | S | S | R | R | R |
E11 | R | R | R | S | R | I |
E16 | R | R | R | S | R | R |
E20 | R | R | R | I | R | R |
E23 | R | R | R | R | R | R |
E26 | R | R | R | R | R | R |
E28 | R | S | S | R | I | S |
E29 | R | R | S | R | R | I |
E30 | R | S | R | R | R | R |
E34 | R | S | I | S | R | R |
E36 | R | R | R | R | R | S |
E40 | R | S | I | S | R | R |
E44 | R | I | R | R | I | R |
E45 | R | R | R | R | R | R |
E46 | S | S | R | S | S | S |
E49 | R | R | R | R | R | R |
E51 | R | S | R | R | S | S |
E52 | R | R | S | R | R | R |
E55 | R | R | R | I | R | R |
E56 | S | S | R | R | I | S |
E58 | S | S | S | R | S | S |
E59 | R | S | R | I | R | R |
E60 | S | S | I | S | S | S |
Controls | R | R | R | R | R | R |
| [1] | B. J. Strasser and T. Schlich, “A history of the medical mask and the rise of throwaway culture,” Lancet, vol. 396, no. 10243, pp. 19–20, Jul. 2020, |
| [2] | Q. X. Ma, H. Shan, H. L. Zhang, G. M. Li, R. M. Yang, and J. M. Chen, “Potential utilities of mask-wearing and instant hand hygiene for fighting SARS-CoV-2,” J. Med. Virol., vol. 92, no. 9, pp. 1567–1571, Sep. 2020, |
| [3] | O. Aydin, B. Emon, S. Cheng, L. Hong, L. P. Chamorro, and M. T. A. Saif, “Performance of fabrics for home-made masks against the spread of COVID-19 through droplets: A quantitative mechanistic study,” Extrem. Mech. Lett., vol. 40, p. 100924, Oct. 2020, |
| [4] | R. N. Mariyati, V. M. L. Toruan, Y. Hutahaean, F. Sihotang, and A. Kartini, “Skin Reactions Due To the Use of Personal Protective Equipment on Healthcare Workers in Covid-19 Pandemic,” J. Ilmu Kesehat., vol. 10, no. 2, pp. 114–121, Dec. 2022, |
| [5] | C. C. I. Foo, A. T. J. Goon, Y. H. Leow, and C. L. Goh, “Adverse skin reactions to personal protective equipment against severe acute respiratory syndrome - A descriptive study in Singapore,” Contact Dermatitis, vol. 55, no. 5, pp. 291–294, Nov. 2006, |
| [6] | M. Yousefimashouf et al., “Evaluation of the bacterial contamination of face masks worn by personnel in a center of COVID 19 hospitalized patients: A cross-sectional study,” New Microbes New Infect., vol. 52, p. 101090, Mar. 2023, |
| [7] |
C. Callewaert, E. De Maeseneire, F. M. Kerckhof, A. Verliefde, T. Van de Wiele, and N. Boon, “Microbial odor profile of polyester and cotton clothes after a fitness session,” Appl. Environ. Microbiol., vol. 80, no. 21, pp. 6611–6619, 2014,
https://doi.org/10.1128/AEM.01422-14/SUPPL_FILE/ZAM999105743SO1.PDF |
| [8] | S. Bin Jeong, K. J. Heo, H. S. Ko, J. P. Ahn, S. B. Lee, and J. H. Jung, “Evaluation of survival rates of airborne microorganisms on the filter layers of commercial face masks,” Indoor Air, vol. 31, no. 4, pp. 1134–1143, Jul. 2021, |
| [9] | P. Luksamijarulkul, N. Aiempradit, and P. Vatanasomboon, “Microbial contamination on used surgical masks among hospital personnel and microbial air quality in their working wards: A hospital in Bangkok,” Oman Med. J., vol. 29, no. 5, pp. 346–350, 2014, |
| [10] | L. Delanghe et al., “Cotton and Surgical Face Masks in Community Settings: Bacterial Contamination and Face Mask Hygiene,” Front. Med., vol. 8, p. 732047, Sep. 2021, |
| [11] | E. J. Lee et al., “Microbial Contamination according to the Numbers of Mask Worn in the Community,” Biomed. Sci. Lett., vol. 28, no. 4, pp. 317–321, Dec. 2022, |
| [12] | A. M. Park et al., “Bacterial and fungal isolation from face masks under the COVID-19 pandemic,” Sci. Rep., vol. 12, no. 1, pp. 1–11, Jul. 2022, |
| [13] | T. Greenhalgh, M. B. Schmid, T. Czypionka, D. Bassler, and L. Gruer, “Face masks for the public during the covid-19 crisis,” BMJ, vol. 369, Apr. 2020, |
| [14] | E. C. Bragginton and L. J. V. Piddock, “UK and European Union public and charitable funding from 2008 to 2013 for bacteriology and antibiotic research in the UK: An observational study,” Lancet Infect. Dis., vol. 14, no. 9, pp. 857–868, Sep. 2014, |
| [15] | J. Slots and H. Slots, “Bacterial and viral pathogens in saliva: disease relationship and infectious risk,” Periodontol. 2000, vol. 55, no. 1, p. 48, Feb. 2011, |
| [16] | N. A. Hasan, B. A. Young, A. T. Minard-Smith, K. Saeed, and H. Li, “Microbial community profiling of human saliva using shotgun metagenomic sequencing (PLoS ONE (2014) 9, 5, (e97699) PLoS One, vol. 9, no. 8, Aug. 2014, |
| [17] | E. J. Derbyshire and P. C. Calder, “Respiratory Tract Infections and Antibiotic Resistance: A Protective Role for Vitamin D?,” Front. Nutr., vol. 8, p. 652469, Mar. 2021, |
| [18] |
CDC, “2019 Antibiotic Resistance Threats Report | CDC,” Center for Disease Control, 2021.
https://www.cdc.gov/drugresistance//biggest-threats.html (accessed Mar. 28, 2024). |
| [19] | K. Adeleye and A. Onifade, “Antibiotic sensitivity profile of microorganisms isolated from used and unused nose masks,” Microbes Infect. Dis., vol. 0, no. 0, pp. 0–0, Nov. 2023, |
| [20] | J. L. Scheid, S. P. Lupien, G. S. Ford, and S. L. West, “Commentary: Physiological and psychological impact of face mask usage during the covid-19 pandemic,” Int. J. Environ. Res. Public Health, vol. 17, no. 18, pp. 1–12, Sep. 2020, |
| [21] | E. N. Kariuki, Z. W. Ng’ang’a, and P. Wanzala, “Bacteriological Contamination Of Street Foods Among Street Food Vendors In Githurai And Gikomba Markets- Nairobi County,” Int. J. Innov. Res. Adv. Stud., vol. 4, no. 1, pp. 337–346, 2017, Accessed: Mar. 20, 2024. [Online]. Available: |
| [22] | B. D. Jett, K. L. Hatter, M. M. Huycke, and M. S. Gilmore, “Simplified agar plate method for quantifying viable bacteria,” Biotechniques, vol. 23, no. 4, pp. 648–650, Aug. 1997, |
| [23] | S. Atashpaz et al., “A robust universal method for extraction of genomic DNA from bacterial species.,” Mikrobiologiia, vol. 79, no. 4, pp. 562–566, Aug. 2010, |
| [24] | A. King and D. F. J. Brown, “Quality assurance of antimicrobial susceptibility testing by disc diffusion,” J. Antimicrob. Chemother., vol. 48, no. SUPPL. 1, pp. 71–76, Jul. 2001, |
| [25] | H. M. Park, “Comparing Group Means: T-tests and One-way ANOVA Using Stata, SAS, R, and SPSS.” 2009. Accessed: Mar. 21, 2024. [Online]. Available: |
| [26] | A. Nanda, D. B. B. Mohapatra, A. P. K. Mahapatra, A. P. K. Mahapatra, and A. P. K. Mahapatra, “Multiple comparison test by Tukey’s honestly significant difference (HSD): Do the confident level control type I error,” Int. J. Stat. Appl. Math., vol. 6, no. 1, pp. 59–65, 2021, |
| [27] | M. Martí, J. Gisbert-Paya, M. Á. Bonet-Aracil, P. Jovančić, M. J. Lis, and L. Coderch, “Increased comfort of polyester fabrics,” Polymers (Basel)., vol. 13, no. 17, p. 3010, Sep. 2021, |
| [28] | P. Gupta, N. Bairagi, R. Priyadarshini, A. Singh, D. Chauhan, and D. Gupta, “Bacterial contamination of nurses’ white coats made from polyester and polyester cotton blend fabrics,” J. Hosp. Infect., vol. 94, no. 1, pp. 92–94, Sep. 2016, |
| [29] | S. Furnaz et al., “Knowledge, attitude and practice of wearing mask in the population presenting to tertiary hospitals in a developing country,” PLoS One, vol. 17, no. 3, p. e0265328, Mar. 2022, |
| [30] |
G. H. Carpenter, “Salivary Factors that Maintain the Normal Oral Commensal Microflora,” J. Dent. Res., vol. 99, no. 6, pp. 644–649, Jun. 2020,
https://doi.org/10.1177/0022034520915486/ASSET/IMAGES/LARGE/10.1177_0022034520915486-FIG3.JPEG |
| [31] | D. Gupta, “Living with in-mask micro-climate,” Med. Hypotheses, vol. 144, p. 110010, Nov. 2020, |
| [32] | J. Marín-Nieto, C. Reino-Perez, G. Santillana-Cernuda, J. M. Díaz-Bernal, R. Luque-Aranda, and I. García-Basterra, “FACE MASK CONTAMINATION during COVID-19 PANDEMIA. A STUDY on PATIENTS RECEIVING INTRAVITREAL INJECTIONS,” Retina, vol. 41, no. 11, pp. 2215–2220, Nov. 2021, |
| [33] | X. Guan et al., “Prolonged Use of Surgical Masks and Respirators Affects the Protection and Comfort for Healthcare Workers,” Materials (Basel)., vol. 15, no. 22, p. 7918, Nov. 2022, |
| [34] | T. Onuoha et al., “Isolation of Bacteria from Constantly used Face Masks of Some Novena University Students,” 2022. |
| [35] | M. P. Gund et al., “Detection of viable oral bacteria of the patient on the surgical mask of dentists,” BDJ Open, 2024. |
| [36] | P. K. Purushothaman, E. Priyangha, and R. Vaidhyswaran, “Effects of Prolonged Use of Facemask on Healthcare Workers in Tertiary Care Hospital During COVID-19 Pandemic,” Indian J. Otolaryngol. Head Neck Surg., vol. 73, no. 1, pp. 59–65, Mar. 2021, |
| [37] | R. Endriani, E. Rafni, A. Bet, H. F. Nabila, M. P. Berlianti, and D. A. Alhadi, “The Bacteria on Saliva of Diabetes Mellitus Patients in Arifin Achmad General Hospital, Riau Province,” Nusant. Sci. Technol. Proc., pp. 49–52, Dec. 2022, |
| [38] | E. K. Allen et al., “Bacteria in the nose of young adults during wellness and rhinovirus colds: Detection by culture and microarray methods in 100 nasal lavage specimens,” Int. Forum Allergy Rhinol., vol. 3, no. 9, pp. 731–739, Sep. 2013, |
| [39] | S. Buerger, “The Skin and Oral Microbiome: An Examination of Overlap and Potential Interactions between Microbiome Communities,” Ski. Microbiome Handb. From Basic Res. to Prod. Dev., pp. 45–57, Jan. 2020, |
| [40] | S. B. DeVore, T. Gonzalez, M. G. Sherenian, A. B. Herr, and G. K. Khurana Hershey, “On the surface: Skin microbial exposure contributes to allergic disease,” Ann. Allergy, Asthma Immunol., vol. 125, no. 6, pp. 628–638, Dec. 2020, |
| [41] | J. R. Adalbert, K. Varshney, R. Tobin, and R. Pajaro, “Clinical outcomes in patients co-infected with COVID-19 and Staphylococcus aureus: a scoping review,” BMC Infect. Dis., vol. 21, no. 1, pp. 1–17, Dec. 2021, |
| [42] | U. V. John and J. Carvalho, “Enterococcus: Review of its physiology, pathogenesis, diseases and the challenges it poses for clinical microbiology,” Front. Biol. (Beijing)., vol. 6, no. 5, pp. 357–366, Oct. 2011, |
| [43] | D. Reynolds and M. Kollef, “The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update,” Drugs, vol. 81, no. 18, pp. 2117–2131, Nov. 2021, |
| [44] | I. S. M. Alhashimi, R. A. Abdul-Jabbar, and S. K. Issa, “The ability of Acinetobacter haemolyticus isolated from soil removes heavy elements (Cd, Pb, Zn, Ni),” Mater. Today Proc., vol. 80, pp. 2448–2457, Jan. 2023, |
| [45] | R. Al-Zakhari, M. Suhail, B. Ataallah, S. Aljammali, and A. Grigos, “Rare but Fatal Case of Cavitary Pneumonia Caused by Alcaligenes Faecalis,” Cureus, vol. 3, 2020, |
| [46] | L. Walsh, S. A. Clark, J. P. Derrick, and R. Borrow, “Beyond the usual suspects: Reviewing infections caused by typically-commensal Neisseria species,” J. Infect., vol. 87, no. 6, pp. 479–489, Dec. 2023, |
| [47] | S. M. Jacobsen and M. E. Shirtliff, “Proteus mirabilis biofilms and catheter-associated urinary tract infections,” Virulence, vol. 2, no. 5, pp. 460–465, 2011, |
| [48] | M. Ehling-Schulz, D. Lereclus, and T. M. Koehler, “ The Bacillus cereus Group: Bacillus Species with Pathogenic Potential,” Microbiol. Spectr., vol. 7, no. 3, May 2019, |
| [49] | A. Bravo, S. Likitvivatanavong, S. S. Gill, and M. Soberón, “Bacillus thuringiensis: A story of a successful bioinsecticide,” Insect Biochem. Mol. Biol., vol. 41, no. 7, pp. 423–431, Jul. 2011, |
| [50] | Y. Bel, J. Ferré, and P. Hernández-Martínez, “Bacillus thuringiensis Toxins: Functional Characterization and Mechanism of Action,” Toxins 2020, Vol. 12, Page 785, vol. 12, no. 12, p. 785, Dec. 2020, |
| [51] | C. Garcia Hejl, N. Sanmartin, T. Samson, C. Soler, and J. L. Koeck, “Sinusite maxillaire due à Bacillus licheniformis: à propos d’un cas survenu à Djibouti,” Med. Sante Trop., vol. 25, no. 2, pp. 220–221, Apr. 2015, |
| [52] | D. M. P. De Oliveira et al., “Antimicrobial resistance in ESKAPE pathogens,” Clin. Microbiol. Rev., vol. 33, no. 3, 2020, |
APA Style
Bitolitoli, E. N., Mutai, M. J., Njeru, E. M., Oyore, J. P., Neondo, J., et al. (2025). Bacteria Contamination and Antibiotic Resistance Profile on Worn Reusable Facemasks in Githurai Market, Kenya. International Journal of Food Science and Biotechnology, 10(2), 33-48. https://doi.org/10.11648/j.ijfsb.20251002.12
ACS Style
Bitolitoli, E. N.; Mutai, M. J.; Njeru, E. M.; Oyore, J. P.; Neondo, J., et al. Bacteria Contamination and Antibiotic Resistance Profile on Worn Reusable Facemasks in Githurai Market, Kenya. Int. J. Food Sci. Biotechnol. 2025, 10(2), 33-48. doi: 10.11648/j.ijfsb.20251002.12
@article{10.11648/j.ijfsb.20251002.12,
author = {Esther Nabwile Bitolitoli and Mourine Jerono Mutai and Ezekiel Mugendi Njeru and John Paul Oyore and Johnstone Neondo and Kennedy Awuor and Daniel Wambiri Muthee and Stephen Super Barasa and Sammy Letema and Richard Okoth Oduor},
title = {Bacteria Contamination and Antibiotic Resistance Profile on Worn Reusable Facemasks in Githurai Market, Kenya
},
journal = {International Journal of Food Science and Biotechnology},
volume = {10},
number = {2},
pages = {33-48},
doi = {10.11648/j.ijfsb.20251002.12},
url = {https://doi.org/10.11648/j.ijfsb.20251002.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijfsb.20251002.12},
abstract = {Facemasks have been widely used in the theater to prevent surgical site infections as well as in hospital wards to prevent infectious infections. Also masks have been recommended to be used by the public during pandemics of respiratory infectious diseases. However, the prolonged use of these masks may have inadvertently harbored unseen dangers by serving as reservoirs for bacteria contamination and antibiotic resistant bacteria when in use. The objective of this study was to quantify bacteria as well as isolate, characterize and determine the antibiotic profiles of bacteria from worn reusable facemasks. Before masks were worn, the participants’ mouth, nose and facial skin were swabbed with sterile transport media. Thereafter, the masks were worn at two-, four- and six-hour intervals. Afterwards, bacteria on worn masks, mouth, nose and skin were identified via morphological, biochemical and molecular methods. Antibacterial susceptibility was determined via the Kirby–Bauer method. The results of this study revealed bacterial colony forming units were significantly higher in the polycotton masks (4.30×102 CFUs) than in the cotton masks (3.38×102 CFUs). Colony forming units increased with extended mask use from two, four, and six hours respectively (7.61×101 CFUs, 1.28×102 CFUs, 4.26×102 CFUs). Also, inside part of the masks had significantly high colony forming units than outside of the masks. The isolated bacteria were from different genera including Bacillus, Staphylococcus, Enterococcus, Pseudomonas, Stenotrophomonas, Acinetobacter, Neisseria, Proteus, Klebsiella and Enterobacter. From the antibiotic resistance profiles, with 85.2% of isolates classified as resistant, Ampicillin had the highest resistance rate. Spectinomycin and Amoxicillin both showed 66.7% resistance, whereas Cefotaxime showed 63.0% resistance. Streptomycin showed the highest susceptibility (48.1%), suggesting comparatively improved efficiency against the isolates. The study findings provides bacteriological insights risks associated with facemask upon use as well as antibiotic resistance.
},
year = {2025}
}
TY - JOUR T1 - Bacteria Contamination and Antibiotic Resistance Profile on Worn Reusable Facemasks in Githurai Market, Kenya AU - Esther Nabwile Bitolitoli AU - Mourine Jerono Mutai AU - Ezekiel Mugendi Njeru AU - John Paul Oyore AU - Johnstone Neondo AU - Kennedy Awuor AU - Daniel Wambiri Muthee AU - Stephen Super Barasa AU - Sammy Letema AU - Richard Okoth Oduor Y1 - 2025/06/25 PY - 2025 N1 - https://doi.org/10.11648/j.ijfsb.20251002.12 DO - 10.11648/j.ijfsb.20251002.12 T2 - International Journal of Food Science and Biotechnology JF - International Journal of Food Science and Biotechnology JO - International Journal of Food Science and Biotechnology SP - 33 EP - 48 PB - Science Publishing Group SN - 2578-9643 UR - https://doi.org/10.11648/j.ijfsb.20251002.12 AB - Facemasks have been widely used in the theater to prevent surgical site infections as well as in hospital wards to prevent infectious infections. Also masks have been recommended to be used by the public during pandemics of respiratory infectious diseases. However, the prolonged use of these masks may have inadvertently harbored unseen dangers by serving as reservoirs for bacteria contamination and antibiotic resistant bacteria when in use. The objective of this study was to quantify bacteria as well as isolate, characterize and determine the antibiotic profiles of bacteria from worn reusable facemasks. Before masks were worn, the participants’ mouth, nose and facial skin were swabbed with sterile transport media. Thereafter, the masks were worn at two-, four- and six-hour intervals. Afterwards, bacteria on worn masks, mouth, nose and skin were identified via morphological, biochemical and molecular methods. Antibacterial susceptibility was determined via the Kirby–Bauer method. The results of this study revealed bacterial colony forming units were significantly higher in the polycotton masks (4.30×102 CFUs) than in the cotton masks (3.38×102 CFUs). Colony forming units increased with extended mask use from two, four, and six hours respectively (7.61×101 CFUs, 1.28×102 CFUs, 4.26×102 CFUs). Also, inside part of the masks had significantly high colony forming units than outside of the masks. The isolated bacteria were from different genera including Bacillus, Staphylococcus, Enterococcus, Pseudomonas, Stenotrophomonas, Acinetobacter, Neisseria, Proteus, Klebsiella and Enterobacter. From the antibiotic resistance profiles, with 85.2% of isolates classified as resistant, Ampicillin had the highest resistance rate. Spectinomycin and Amoxicillin both showed 66.7% resistance, whereas Cefotaxime showed 63.0% resistance. Streptomycin showed the highest susceptibility (48.1%), suggesting comparatively improved efficiency against the isolates. The study findings provides bacteriological insights risks associated with facemask upon use as well as antibiotic resistance. VL - 10 IS - 2 ER -
Biochemistry, Microbiology and Biotechnology, Kenyatta University School of Pure and Applied Sciences, Nairobi, Kenya
Biochemistry, Microbiology and Biotechnology, Kenyatta University School of Pure and Applied Sciences, Nairobi, Kenya
Biochemistry, Microbiology and Biotechnology, Kenyatta University School of Pure and Applied Sciences, Nairobi, Kenya
Family Medicine, Community Health and Epidemiology, Kenyatta University School of Health Sciences, Nairobi, Kenya
Institute of Biotechnology Research, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
Mathematics, Kenyatta University School of Pure and Applied Sciences, Nairobi, Kenya
Computing and Information Science, Kenyatta University, Nairobi, Kenya
Chemistry and Biochemistry, University of Eldoret, Eldoret, Kenya
Spatial and Environmental Planning, Kenyatta University, Nairobi, Kenya0009-0007-9624-6580
Biochemistry, Microbiology and Biotechnology, Kenyatta University School of Pure and Applied Sciences, Nairobi, Kenya
Information