The increasing commoditization of traditional biostimulants has driven the need for more precise and targeted solutions in climate change scenarios. Single Biostimulant Molecules (SBM) represent a novel approach that departs from conventional multi-compound formulations, which often contain undefined mixtures of bioactive components, triggering multiple responses. Instead, SBMs focus on a specific bioactive molecule with a defined mode of action, ensuring greater consistency and predictability in plant responses under field conditions. This study evaluated Carrabiitol®, an oligosaccharide polyol single biostimulant molecule, for alleviating stress response in tomato (Solanum lycopersicum cv. ‘Arka Rakshak’). Plants were subjected to water deficit, high temperature, and salinity stress during the flowering stages. Treatment involved seed priming and application of Carrabiitol® (3ml/L) on foliage during vegetative and pre-flowering stages. Plants were evaluated for their Plant height, Number of fruits per plant, Total Soluble sugars, Total Acidity, Lycopene content, Carotenoids content and fruit yield. Experiments were arranged in a complete block randomized design with four replications. Results showed recovery in plant height and marketable yield with significant improvement in the tomato lycopene content that increased by more than 76% under water deficit stress and 164% under salinity stress when treated with Carrabiitol® @ 3ml/L (T6) compared to untreated plants (p ≤ 0.05). Total carotenoids improved by up to 89% (p ≤ 0.05). Overall, study illustrated that application of Single Biostimulant Molecule (SBM), Carrabiitol® was effective in improving the qualitative trait of tomatoes by alleviating applied abiotic stresses.
| Published in | Journal of Plant Sciences (Volume 14, Issue 2) |
| DOI | 10.11648/j.jps.20261402.14 |
| Page(s) | 104-114 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Tomato, Abiotic Stress, Carrabiitol®, Climate Change, Single Biostimulant Molecule (SBM)
Treatment | Details |
|---|---|
T1 | Plant raised from seeds without Carrabiitol |
T2 | Plants raised from seeds treated with 3 ml/L Carrabiitol + Foliar spray at 2-3 leaf stage |
T3 | Plants raised from seeds treated with 3 ml/L Carrabiitol + Foliar spray at 2-3 leaf stage+ Foliar spray at pre-flowering stage |
T4 | Plant raised from seeds without Carrabiitol + Stress at flowering stage |
T5 | Plants raised from seeds treated with 3 ml/L Carrabiitol + Foliar spray at 2-3 leaf stage + Stress at flowering stage |
T6 | Plants raised from seeds treated with 3 ml/L Carrabiitol + Foliar spray at 2-3 leaf stage+ Foliar spray at pre-flowering stage + Stress at flowering stage |
T7 | Plants raised from seeds + Foliar spray @ 5m/L commercial control (Mixture of seaweed & Protein hydrolysate) at 2-3 leaf stage+ Foliar spray at pre-flowering stage + Stress at flowering stage |
Treatment | Date |
|---|---|
Soaking of seeds in 3ml/L solution | 21.08.2022 |
Date of sowing in protrays | 22.08.2022 |
Transplant of seedlings to pots | 23.09.2022 |
Booster dose at vegetative stage | 06.10.2022 |
Booster dose at pre-flowering stage | 22.10.2022 |
Drought stress imposition | 06.11.2022 to 11.11.2022 |
High temperature stress imposition (Trial 1) | 02.11.2022 to 05.11.2022 |
High temperature stress imposition (Trial 2) | 20.08.2024 to 23.08.2024 |
Salinity stress imposition | 07.11.2022 to 15.11.2022 |
Date of final harvest | 10.01.2023 |
SBM | Single Biostimulant Molecule |
LSD | Least Significant Differences |
ROS | Reactive Oxygen Species |
| [1] | Alexandratos, N., Bruinsma, J., 2012. World agriculture towards 2030/2050: the 2012 revision. ESA Working Papers 12-03. |
| [2] |
UN, 2017. World population projected to reach 9.8 billion in 2050, and 11.2 billion in 2100 [WWW Document]. U. N. URL
https://www.un.org/en/desa/world-population-projected-reach-98-billion-2050-and-112-billion-2100 (accessed 7.9.25). |
| [3] | The State of Food Security and Nutrition in the World 2022, 2022. FAO. |
| [4] | Kürklü, A., Pearson, S., Felek, T., 2025. Climate change impacts on tomato production in high-tech soilless greenhouses in Türki̇ye. BMC Plant Biol. 25, 339. |
| [5] | Ashraf, M., Harris, P. J. C., 2004. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 166, 3–16. |
| [6] | Atta, K., Mondal, Saptarshi, Gorai, S., Singh, A. P., Kumari, A., Ghosh, T., Roy, A., Hembram, S., Gaikwad, D. J., Mondal, Subhasis, Bhattacharya, S., Jha, U. C., Jespersen, D., 2023. Impacts of salinity stress on crop plants: improving salt tolerance through genetic and molecular dissection. Front. Plant Sci. 14, 1241736. |
| [7] | Nishiyama, R., Watanabe, Y., Leyva-Gonzalez, M. A., Van Ha, C., Fujita, Y., Tanaka, M., Seki, M., Yamaguchi-Shinozaki, K., Shinozaki, K., Herrera-Estrella, L., Tran, L.-S. P., 2013. Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response. Proc. Natl. Acad. Sci. 110, 4840–4845. |
| [8] | Osakabe, Y., Osakabe, K., Shinozaki, K., Tran, L.-S. P., 2014. Response of plants to water stress. Front. Plant Sci. 5, 00086. |
| [9] | Singh, P., Choudhary, K. K., Chaudhary, N., Gupta, S., Sahu, M., Tejaswini, B., Sarkar, S., 2022. Salt stress resilience in plants mediated through osmolyte accumulation and its crosstalk mechanism with phytohormones. Front. Plant Sci. 13, 1006617. |
| [10] | du Jardin, P., 2015. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic., Biostimulants in Horticulture 196, 3–14. |
| [11] | EL Boukhari, M. E. M., Barakate, M., Bouhia, Y., Lyamlouli, K., 2020. Trends in Seaweed Extract Based Biostimulants: Manufacturing Process and Beneficial Effect on Soil-Plant Systems. Plants 9, 359. |
| [12] | Colla, G., Rouphael, Y., 2015. Biostimulants in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 1–2. |
| [13] | Pichyangkura, R., Chadchawan, S., 2015. Biostimulant activity of chitosan in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 49–65. |
| [14] | Canellas, L. P., Olivares, F. L., Aguiar, N. O., Jones, D. L., Nebbioso, A., Mazzei, P., Piccolo, A., 2015. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 15–27. |
| [15] | Colla, G., Nardi, S., Cardarelli, M., Ertani, A., Lucini, L., Canaguier, R., Rouphael, Y., 2015. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 28–38. |
| [16] | Gomez-Casati, Iglesias, A., 2002. ADP-glucose pyrophosphorylase from wheat endosperm. Purification and characterization of an enzyme with novel regulatory properties. Planta 214, 428–434. |
| [17] | Battacharyya, D., Babgohari, M. Z., Rathor, P., Prithiviraj, B., 2015. Seaweed extracts as biostimulants in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 39–48. |
| [18] | Savvas, D., Ntatsi, G., 2015. Biostimulant activity of silicon in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 66–81. |
| [19] | Rouphael, Y., Franken, P., Schneider, C., Schwarz, D., Giovannetti, M., Agnolucci, M., Pascale, S. D., Bonini, P., Colla, G., 2015. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic., Biostimulants in Horticulture 196, 91–108. |
| [20] | Ruzzi, M., Aroca, R., 2015. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic., Biostimulants in Horticulture 196, 124–134. |
| [21] | López-Bucio, J., Pelagio-Flores, R., Herrera-Estrella, A., 2015. Trichoderma as biostimulant: exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic., Biostimulants in Horticulture 196, 109–123. |
| [22] | MM, E. S., Ismail, M. M. and Hamouda, M. M. (2016). Influence of some brown seaweed extracts on germination and cytological responses of Trigonella foenum-graecum L. BioTechnology: An Indian Journal. 12: 1-12. |
| [23] | Li, J., Lardon, R., Mangelinckx, S., & Geelen, D. (2024). A practical guide to the discovery of biomolecules with biostimulant activity. Journal of Experimental Botany, 75(13), 3797–3817. |
| [24] | Mashabela, M. D., Terefe, T., Kerchev, P., Sitole, L., & Mhlongo, M. I. (2025). Next-generation biostimulants: molecular insights, digital integration, and regulatory frameworks for sustainable agriculture. Frontiers in Plant Science, 16, 1710899. |
| [25] | Patel, F. Y., Upreti, K. K., Laxman, R. H., Shah, N. J., 2024. Carrabiitol®, a Novel Oligosaccharide Polyol Composition, Mitigates the Impact of Flooding, Drought, Salinity, and High Temperature in Tomato. Biology 13, 356. |
| [26] | Zahra, N., Raza, Z. A., Mahmood, S., 2020. Effect of Salinity Stress on Various Growth and Physiological Attributes of Two Contrasting Maize Genotypes. Braz. Arch. Biol. Technol. 63, e20200072. |
| [27] | Roșca, M., Mihalache, G., Stoleru, V., 2023. Tomato responses to salinity stress: From morphological traits to genetic changes. Front. Plant Sci. 14, 1118383. |
| [28] | Hasegawa, P. M., 2013. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot., Sustainable cultivation and exploitation of halophyte crops in a salinizing world 92, 19–31. |
| [29] | Shrivastava, P., Kumar, R., 2015. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 22, 123–131. |
| [30] | He, J., Han, W., Wang, J., Qian, Y., Saito, M., Bai, W., Song, J., Lv, G., 2022. Functions of Oligosaccharides in Improving Tomato Seeding Growth and Chilling Resistance. J. Plant Growth Regul. 41, 535–545. |
| [31] | Campestrini, L. H., Melo, P. S., Peres, L. E. P., Calhelha, R. C., Ferreira, I. C. F. R., Alencar, S. M., 2019. A new variety of purple tomato as a rich source of bioactive carotenoids and its potential health benefits. Heliyon 5, e02831. |
| [32] | Li, N., Wu, X., Zhuang, W., Xia, L., Chen, Y., Wu, C., Rao, Z., Du, L., Zhao, R., Yi, M., Wan, Q., Zhou, Y., 2021. Tomato and lycopene and multiple health outcomes: Umbrella review. Food Chem. 343, 128396. |
| [33] | Zheng, J., Chen, H., Wang, T., Mustafa, G., Liu, L., Wang, Q., Shao, Z., 2023. Quality Improvement of Tomato Fruits by Preharvest Application of Chitosan Oligosaccharide. Horticulturae 9, 300. |
| [34] | Chi, Y., Wang, X., Li, F., Zhang, Z., 2025. The Effects of Alginate Oligosaccharides (AOS) with Degree of Polymerization (DP) 2, 3, 4, 5, 6, and 7 on Tomato Yields. |
APA Style
Patel, F. Y., Veeraiah, R. K., Laxman, R. H., Shah, N. J. (2026). Carrabiitol Mediated Qualitative Enhancement of Tomato Fruit Under Abiotic Stress Conditions. Journal of Plant Sciences, 14(2), 104-114. https://doi.org/10.11648/j.jps.20261402.14
ACS Style
Patel, F. Y.; Veeraiah, R. K.; Laxman, R. H.; Shah, N. J. Carrabiitol Mediated Qualitative Enhancement of Tomato Fruit Under Abiotic Stress Conditions. J. Plant Sci. 2026, 14(2), 104-114. doi: 10.11648/j.jps.20261402.14
@article{10.11648/j.jps.20261402.14,
author = {Femida Yunus Patel and Ramesh Karugahalli Veeraiah and Ramanna Hunashikatti Laxman and Neil Jaykumar Shah},
title = {Carrabiitol Mediated Qualitative Enhancement of Tomato Fruit Under Abiotic Stress Conditions},
journal = {Journal of Plant Sciences},
volume = {14},
number = {2},
pages = {104-114},
doi = {10.11648/j.jps.20261402.14},
url = {https://doi.org/10.11648/j.jps.20261402.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.jps.20261402.14},
abstract = {The increasing commoditization of traditional biostimulants has driven the need for more precise and targeted solutions in climate change scenarios. Single Biostimulant Molecules (SBM) represent a novel approach that departs from conventional multi-compound formulations, which often contain undefined mixtures of bioactive components, triggering multiple responses. Instead, SBMs focus on a specific bioactive molecule with a defined mode of action, ensuring greater consistency and predictability in plant responses under field conditions. This study evaluated Carrabiitol®, an oligosaccharide polyol single biostimulant molecule, for alleviating stress response in tomato (Solanum lycopersicum cv. ‘Arka Rakshak’). Plants were subjected to water deficit, high temperature, and salinity stress during the flowering stages. Treatment involved seed priming and application of Carrabiitol® (3ml/L) on foliage during vegetative and pre-flowering stages. Plants were evaluated for their Plant height, Number of fruits per plant, Total Soluble sugars, Total Acidity, Lycopene content, Carotenoids content and fruit yield. Experiments were arranged in a complete block randomized design with four replications. Results showed recovery in plant height and marketable yield with significant improvement in the tomato lycopene content that increased by more than 76% under water deficit stress and 164% under salinity stress when treated with Carrabiitol® @ 3ml/L (T6) compared to untreated plants (p ≤ 0.05). Total carotenoids improved by up to 89% (p ≤ 0.05). Overall, study illustrated that application of Single Biostimulant Molecule (SBM), Carrabiitol® was effective in improving the qualitative trait of tomatoes by alleviating applied abiotic stresses.},
year = {2026}
}
TY - JOUR T1 - Carrabiitol Mediated Qualitative Enhancement of Tomato Fruit Under Abiotic Stress Conditions AU - Femida Yunus Patel AU - Ramesh Karugahalli Veeraiah AU - Ramanna Hunashikatti Laxman AU - Neil Jaykumar Shah Y1 - 2026/04/30 PY - 2026 N1 - https://doi.org/10.11648/j.jps.20261402.14 DO - 10.11648/j.jps.20261402.14 T2 - Journal of Plant Sciences JF - Journal of Plant Sciences JO - Journal of Plant Sciences SP - 104 EP - 114 PB - Science Publishing Group SN - 2331-0731 UR - https://doi.org/10.11648/j.jps.20261402.14 AB - The increasing commoditization of traditional biostimulants has driven the need for more precise and targeted solutions in climate change scenarios. Single Biostimulant Molecules (SBM) represent a novel approach that departs from conventional multi-compound formulations, which often contain undefined mixtures of bioactive components, triggering multiple responses. Instead, SBMs focus on a specific bioactive molecule with a defined mode of action, ensuring greater consistency and predictability in plant responses under field conditions. This study evaluated Carrabiitol®, an oligosaccharide polyol single biostimulant molecule, for alleviating stress response in tomato (Solanum lycopersicum cv. ‘Arka Rakshak’). Plants were subjected to water deficit, high temperature, and salinity stress during the flowering stages. Treatment involved seed priming and application of Carrabiitol® (3ml/L) on foliage during vegetative and pre-flowering stages. Plants were evaluated for their Plant height, Number of fruits per plant, Total Soluble sugars, Total Acidity, Lycopene content, Carotenoids content and fruit yield. Experiments were arranged in a complete block randomized design with four replications. Results showed recovery in plant height and marketable yield with significant improvement in the tomato lycopene content that increased by more than 76% under water deficit stress and 164% under salinity stress when treated with Carrabiitol® @ 3ml/L (T6) compared to untreated plants (p ≤ 0.05). Total carotenoids improved by up to 89% (p ≤ 0.05). Overall, study illustrated that application of Single Biostimulant Molecule (SBM), Carrabiitol® was effective in improving the qualitative trait of tomatoes by alleviating applied abiotic stresses. VL - 14 IS - 2 ER -
Agri Biochem Research Lab, M/s. Pushpa J. Shah, GIDC Panoli, India
Division of Biosciences, Indian Institute of Horticulture Research-ICAR, Bengaluru, India
Agri Biochem Research Lab, M/s. Pushpa J. Shah, GIDC Panoli, India

Figure 1. Tomato plants (Solanum lycopersicum cv. Arka Rakshak) raised on protrays from seeds pre-treated with 3 mL/L Carrabiitol® solution.

Figure 2. Experimental Set up for Tomato plants (Solanum lycopersicum cv. Arka Rakshak) grown under (A) water deficit stress and salinity stress in rain-out shelter, (B) Plants exhibit the onset of fruit production after recovery from stress imposition and (C) heat stress in growth chamber.
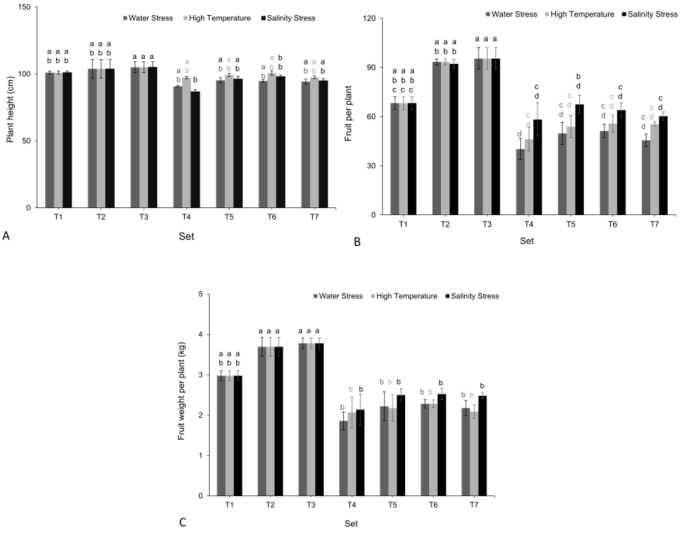
Figure 3. Effect of Carrabiitol® treatments on tomato cultivar 'Arka Rakshak' (A) plant height (cm), (B) number of fruits per plant (no.), and (C) fruit weight per plant (kg) were observed under water deficit, high temperature, and salinity stress conditions. The means not sharing a common letter are significantly different by post hoc least significant difference (LSD) test at 5% level of significance. Data presented are means ± standard errors (p ≤ 0.05, n=4). T1-T7 represent different treatment as described in the M&M section.
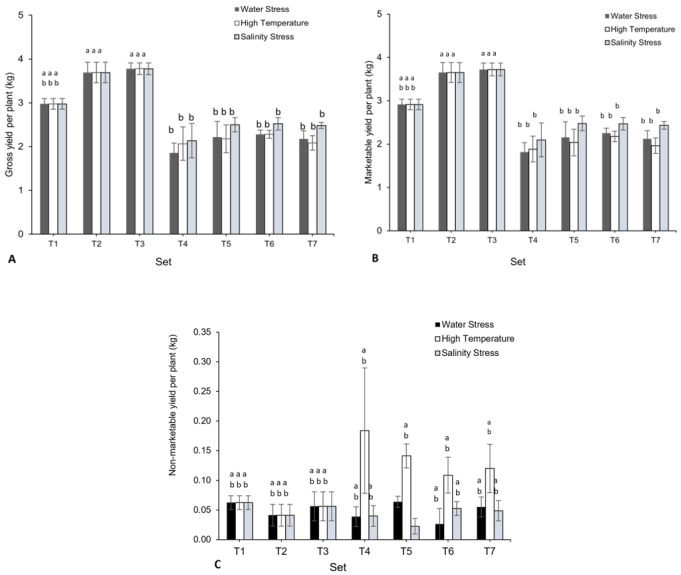
Figure 4. Effect of Carrabiitol® treatments on tomato variety 'Arka Rakshak' (A) Gross yield per plant (kg), (B) Marketable yield per plant (kg), and (C) Non-marketable yield per plant (kg) under water deficit, high temperature, and salinity stress conditions. The means not sharing a common letter are significantly different by post hoc least significant difference (LSD) test at 5% level of significance. Data presented are means ± standard errors (p ≤ 0.05, n=4). T1-T7 represent different treatment as described in the M&M section.
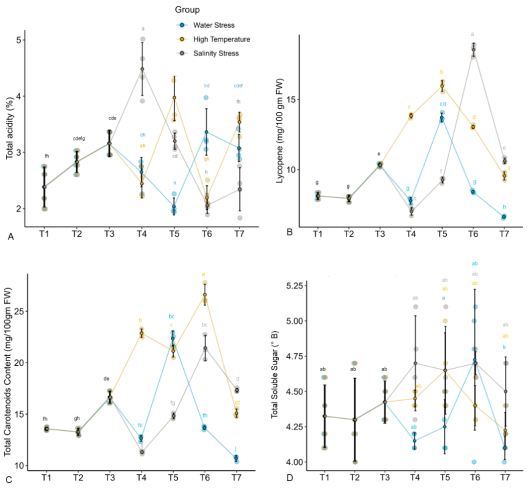
Figure 5. Effect of Carrabiitol® treatments on fruit quality parameters of tomato variety 'Arka Rakshak’. (A) Total acidity (%) (B) Lycopene content (mg/100g FW) (C) Total carotenoid content (mg/100g FW) (D) Total soluble sugar (TSS) content (mg/100g FW) under water deficit, high temperature, and salinity stress conditions. The means not sharing a common letter are significantly different by post hoc least significant difference (LSD) test at 5% level of significance. Data presented are means ± standard errors (p ≤ 0.05, n=4). T1-T7 represent different treatment as described in the M&M section.
Information

